
太陽光は全ての生命にとって重要なエネルギー源です。光合成生物は光合成によって太陽光エネルギーを獲得するために、周囲の環境に合わせて“光のアンテナ”を調節し効果的に光を集めています。今回、基礎生物学研究所および生理学研究所/生命創成探究センターを中心とした研究チームは、ピコプランクトンの一種、オストレオコッカス(プラシノ藻)の光化学系I超複合体の立体構造を決定しました(解像度は2.9Å)。プラシノ藻は水の中に棲む緑藻と、陸地に上がった「植物」の共通の祖先の名残をとどめる原始緑藻です。その光化学系Iは緑藻型と陸上植物型の特徴を兼ね備えていました。さらに、光化学系IIのアンテナの一部も結合した「ステート遷移型」構造であることもわかりました。祖先型光化学系Iがこのようなユニークな構造であることは極めて興味深く、「生物進化に合わせて光化学系がどのような進化を遂げたのか」、逆に「光化学系の進化が生物進化にどう影響したのか」、について今後議論が深まると予想されます。本研究は自然科学研究機構 基礎生物学研究所の石井麻子研究員、キム・ウンチュル助教、皆川純教授、生理学研究所/生命創成探究センター(ExCELLS)の宋致宖特任助教、村田和義特任教授、中国科学院のZhenfeng Liu博士らによる国際共同研究チームによる成果です。
研究成果は英国の総合科学誌eLife(英国時間2023年4月12日付)に掲載されました。
祖先型光化学系Ⅰ超複合体の構造 緑藻型と陸上植物型の特徴を兼ね備え、さらに…
研究の背景
太陽光は全ての生命にとって重要なエネルギー源です。光合成生物は光合成により太陽光エネルギーを獲得しますが、それにはまず光を集めなければなりません。光を集めるにあたり、光合成生物は周囲の環境に合わせて“光のアンテナ”(集光アンテナ複合体; Light Harvesting Complex, LHC)の大きさや性質を調節し効率良く光を集めていることがわかってきました。エネルギー変換を行う装置には光化学系I(PSI)と光化学系II(PSII)の二種類があり、それぞれが光のアンテナを備えています。周囲が暗い時には光のアンテナを大きく広げる必要があります。逆に周囲が明るい時には小さくする必要があります。光の量が急に大きくなったとき、光の色が急に変わった時には瞬時にアンテナの性質が変わることもわかっています(→ 基礎生物学研究所プレスリリース 2016年9月15日「青色光受容体が光合成にブレ―キをかけることを発見 〜青い光が光合成装置を守る〜」)。
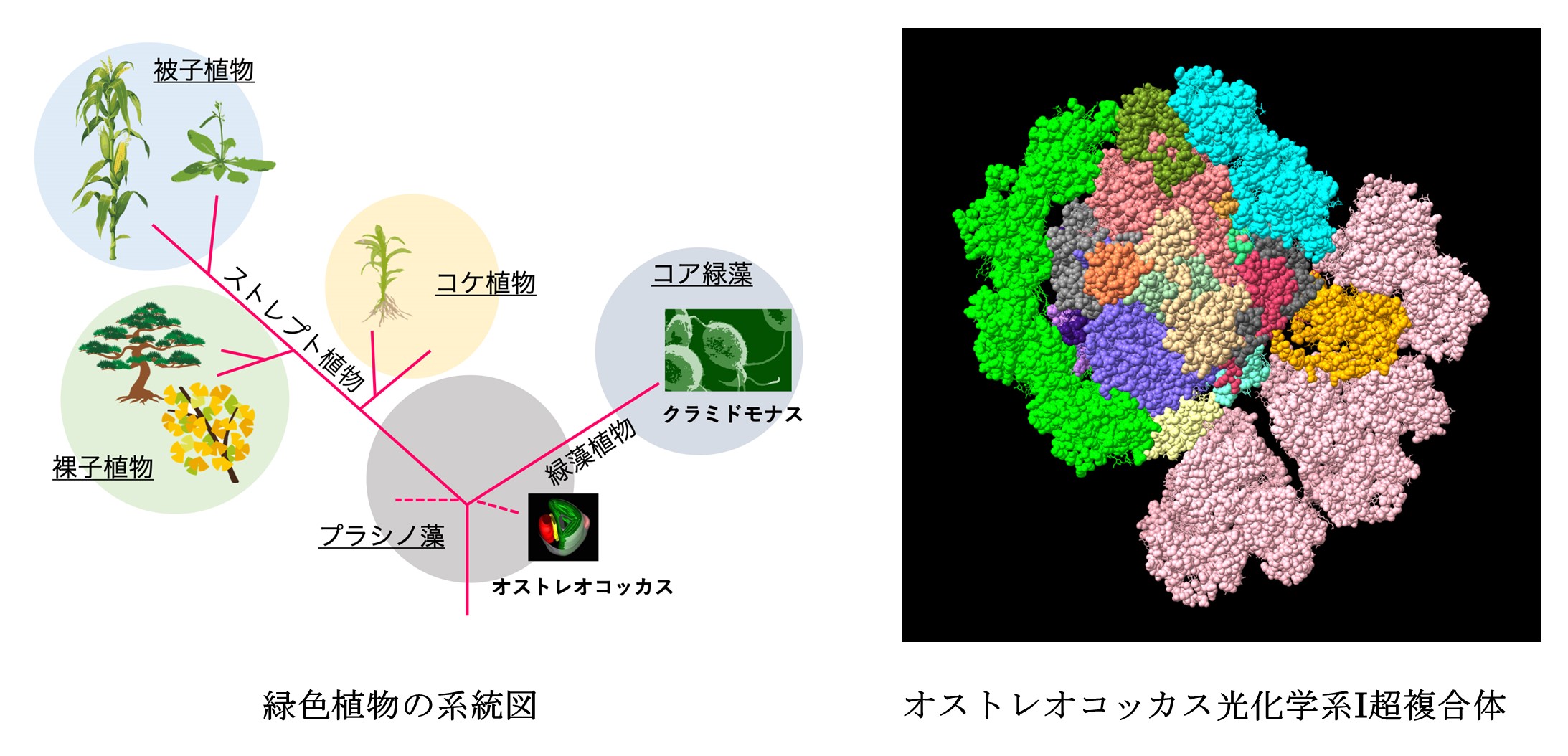
私たちにとって身近な緑色植物は、水の中に棲む「緑藻」と陸に上がる「ストレプト植物」に大きく分類することができます。「緑藻」の中でも後期に出現したコア緑藻や「ストレプト植物」の中でも後期に出現した陸上植物については光合成器官の情報が蓄積されつつあります。例えば最近明らかになったPSIのステート2構造(→ 基礎生物学研究所プレスリリース 2021年7月9日「光合成のステート遷移構造決まる」)の例を見てみると、陸上植物であるトウモロコシの構造と緑藻クラミドモナスの構造には共通点も多いものの、トウモロコシでは移動性三量体LHCが1つ結合しているのに対しクラミドモナスでは2つ結合しているなど、それぞれの生物の生存環境や進化の歴史を反映した違いがあることがわかってきました。では、緑藻と陸上植物が分岐した進化初期に誕生した種(原始緑藻)のPSIはどうだったのでしょうか。今回研究に用いられたのは、そんな最小の真核単細胞藻類(ピコプランクトン)であるプラシノ藻オストレオコッカス(Ostreococcus tauri)です。
研究の成果
研究グループは、オストレオコッカス細胞を弱光環境と強光環境で培養したところ、PSIは弱光環境では“大きな”光アンテナを持ち、強光環境では“小さな”光アンテナを持つことがわかりました(図1)。低解像度の構造解析の結果、“大きな”光アンテナを持つPSIはこれまで報告されたことのないユニークな構造であったため、クライオ電子顕微鏡によってさらに高解像度の構造決定を行いました。
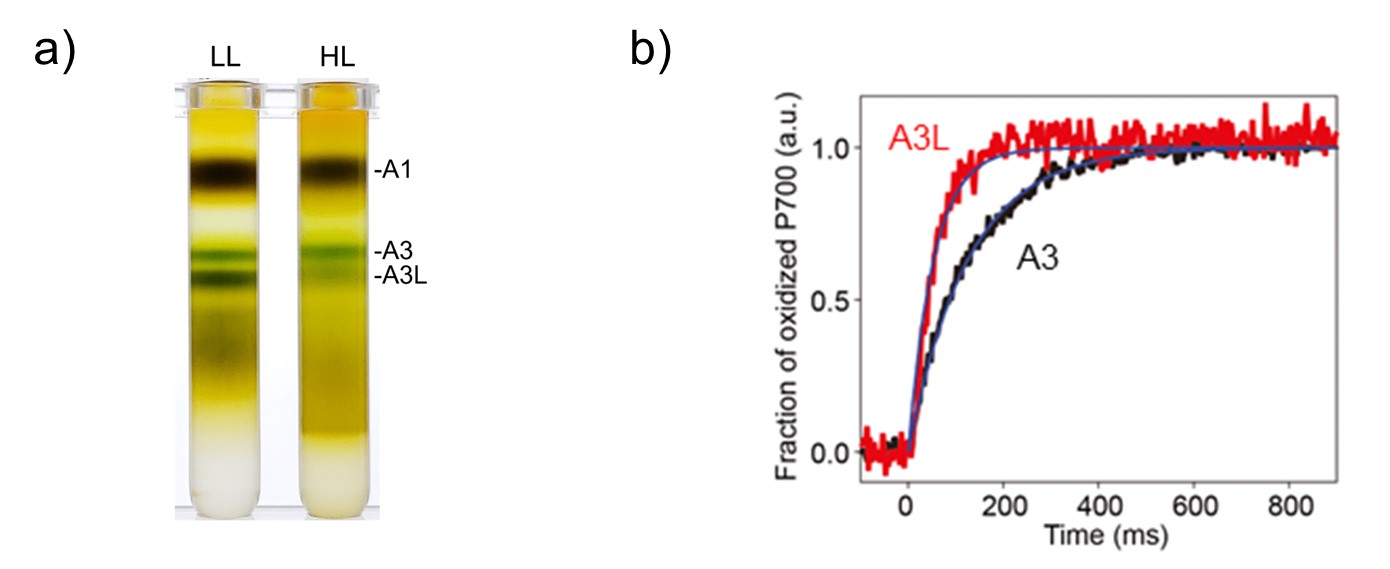 図1:オストレオコッカス光化学系I(PSI)の弱光(LL)条件と強光条件(HL)の比較.
図1:オストレオコッカス光化学系I(PSI)の弱光(LL)条件と強光条件(HL)の比較.
a) ショ糖密度勾配超遠心―チラコイド膜を可溶化し、ショ糖密度勾配超遠心法により大きさに従って分離した。A3画分とA3L画分がPSI超複合体に相当する。LLの時にA3L画分が濃いことがわかる。
b) P700+再還元法によるPSI集光アンテナサイズ解析―PSIのスペシャルペアであるP700にいかに速くエネルギーが伝達されるかで集光アンテナサイズを見積もることができる。A3Lの集光アンテナはA3の約2.2倍の大きさであることが示されている。
こうして決定されたのは、電荷分離が行われるコア複合体の回りを集光アンテナ複合体I(LHCI)とプラシノ藻固有の集光アンテナ複合体Lhcpが取り囲み大きく集光能力を向上させた構造でした。コア複合体の片側に結合した一層の「LHCIベルト」は、陸上植物のPSI超複合体と共通しています(図2:左側、図3)。もう片側のPsaGとPsaHの間の「LHCIヘテロダイマー」は、クラミドモナスのPSI超複合体と共通しています(図2上側、図3)。つまり、オストレオコッカスのPSI超複合体は、祖先型らしく陸上植物と緑藻のPSI超複合体の両方の特徴を兼ね備えていたのです。さらに興味深いことに、第三の側面であるLhca6とPsaKの間には3個の三量体Lhcp(Trimer 1-3)が結合していました(図2右側、図3)。この第三の側面は、緑藻や陸上植物では、ステート遷移時にPSIIから切り離されたLHC三量体が再結合することで知られています(→ 基礎生物学研究所プレスリリース 2021年7月9日)。また、Trimer 2に含まれるLhcp1のN末端スレオニン残基はリン酸化されていることもわかりました。
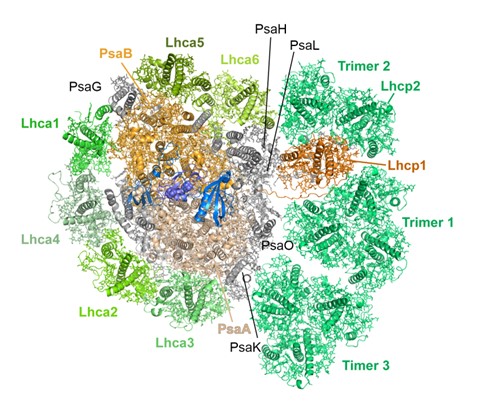 図2:オストレオコッカス光化学系I超複合体(PSI-LHCI-Lhcp)構造.
図2:オストレオコッカス光化学系I超複合体(PSI-LHCI-Lhcp)構造.
葉緑体チラコイド膜鉛直方向ストロマ側から見下ろした全体構造.中央部分(茶)が光化学系I(PSI)コアであり、その周りを以下の集光アンテナが取り囲む構造である。左側に1層の「LHCIベルト」(Lhca1-Lhca4-Lhca2-Lhca3;緑)が結合し、上部には「LHCIハーフベルト」(Lhca5-Lhca6;オリーブ)が、さらに右側に3つのLhcp三量体(Trimer 1-3)が結合している。Trimer 2内のLhcp1のN末端Thr残基はリン酸化されており、PSIコアのPsaLのArg残基と相互作用している。
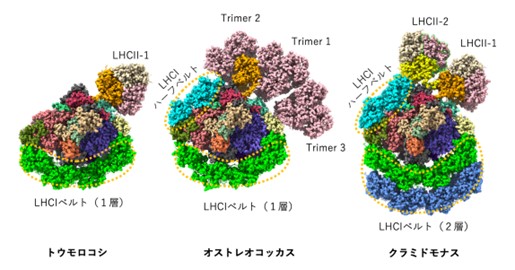 図3:トウモロコシPSI超複合体構造(ステート2)、オストレオコッカスPSI超複合体構造(弱光)、クラミドモナスPSI超複合体構造(ステート2)の比較.
図3:トウモロコシPSI超複合体構造(ステート2)、オストレオコッカスPSI超複合体構造(弱光)、クラミドモナスPSI超複合体構造(ステート2)の比較.
中心に位置するコア複合体にて光化学反応(光エネルギーが電気化学エネルギーに変換される)が行われるが、そこに光エネルギーを供給する集光アンテナの構造がそれぞれの種で異なる。トウモロコシやクラミドモナスの場合、通常時はLHCII-1やLHCII-2などの三量体LHCは結合しておらず、PSIとPSIIの励起バランスを補正する必要が生じた時のみ結合する(ステート2構造)。一方、弱光条件で育てたオストレオコッカスは三量体LHCを3個(Trimer 1-3)結合していた。
ステート遷移とLHCのリン酸化
PSIIとPSIはエネルギー順位では直列に並ぶため、光合成を効率良く行うためには2つの光化学系がバランスよく駆動され葉緑体上の電子移動がスムーズに行われる必要があります。緑藻や陸上植物には、2つの光化学系の励起バランスが崩れてもこれを瞬時に補正するステート遷移(*1)と呼ばれる精緻なしくみが備わっています。最近、ステート遷移の際に重要な「鍵と鍵穴構造」(LHCのリン酸化スレオニン残基とPsaLのアルギニン残基)が明らかになりましたが、陸上植物トウモロコシと緑藻クラミドモナスは共にこの構造を持っています(→ 基礎生物学研究所プレスリリース 2021年7月9日 )。興味深いことに、今回明らかになったオストレオコッカスLhcp1のリン酸化スレオニン残基とそれが結合しているPsaLのアルギニン残基の構造もまた、このステート遷移の「鍵と鍵穴構造」に酷似していたのです(図4)。
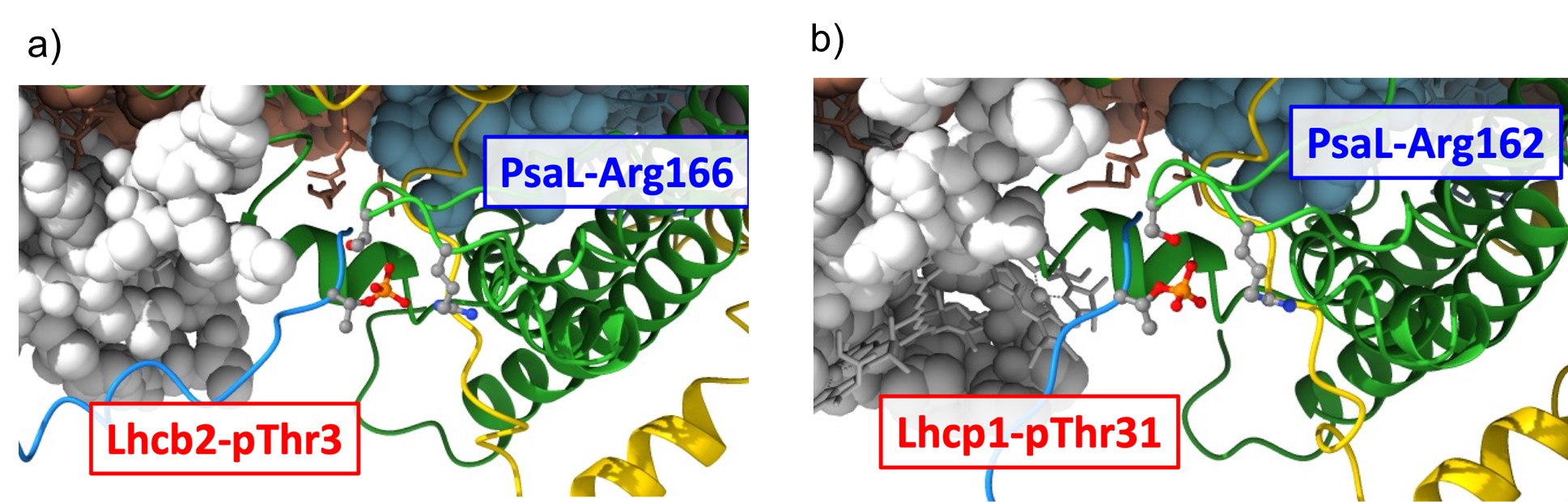 図4:PSI超複合体ステート2構造(トウモロコシ, a)とPSI超複合体弱光構造(オストレオコッカス, b)の“鍵と鍵穴”構造.
図4:PSI超複合体ステート2構造(トウモロコシ, a)とPSI超複合体弱光構造(オストレオコッカス, b)の“鍵と鍵穴”構造.
LHC-N末端Arg-Arg-pThr (リン酸化Thr)とPsaL-Argとの相互作用を示す。LHC(青), PsaL(緑), pThr(赤/黄ball&stick), PsaL-Arg(灰ball&stick)
このことは、オストレオコッカスのPSIに結合している3つの三量体Lhcpはステート遷移の結果PSIと再結合したものであると想像させます。ところが、ことはそう単純ではありません。第一に、このオストレオコッカスは弱光環境で育てられたものであり、PSIIを特に優先的に励起した(ステート2条件)わけではありません。第二に、オストレオコッカスはそもそもステート遷移を起こさない生物である可能性も指摘されています。その理由は、オストレオコッカスのPSIには“レッドクロロフィル”(*2)が存在しないなど、PSIとPSIIに分光学的な差が余りなく、バランス補正の必要はないかもしれないのです。にもかかわらず、今回明らかになったPSI構造はなぜ三量体LHCを“ステート2”座位に結合させているのでしょうか?今後の研究に期待がかかります。
本研究の意義と今後の展開
今回解明されたプラシノ藻オストレオコッカスのPSI超複合体の構造は、基本骨格が植物型と緑藻型の祖先型であるとともに、リン酸化LHC三量体が“ステート2”座位に結合していることを示していました。リン酸化LHCのPSI結合はプラシノ藻が分岐した大昔から行われていたと考えられます。残された謎は、このLHCリン酸化が、のちに登場した緑藻や陸上植物と同様にステート遷移のためのものであるのか?、あるいはPSIのアンテナを長期的に拡大するためのものであるのか?、です。仮に前者であるとすると、オストレオコッカスにはバランス補正をする必要性が見当たらないこととの整合性がつきません。さらなる研究の展開が待たれます。今後の研究によってLHCリン酸化の当初の役割が明らかになると、今日栄華を誇る陸上植物の光合成様式がいかに成立したのか、そして光合成と植物進化はどう関係してきたのかの理解が深まることでしょう。また、これらの知見は、さまざまな環境における光合成デザインへとつながるものと期待されます。
用語解説
*1 : ステート遷移:植物や緑藻のPSIIはPSIと比べて青色光をよく吸収し、一方PSIはPSIIと比べて赤や近赤外光をよく吸収する。このような分光学的特性の違いにより、周囲の光の色が変わったときなど、PSIIとPSIの励起バランスは崩れることがある。そのような時に発動するバランス補正システムがステート遷移である。ステート遷移では、PSIIに結合していた集光アンテナLHCのN末端スレオニン残基(鍵)がリン酸化されてPSIIから解離し、PSIコア複合体(PsaLサブユニットのアルギニン残基、鍵穴)と再結合する。
*2 : レッドクロロフィル:シアノバクテリアから緑藻、陸上植物にいたるまでほとんどの生物のPSIは、通常よりも赤外領域よりの吸収帯(700nm付近)と近赤外領域の蛍光発光帯(715~740nm)を持つ通称“レッドクロロフィル”と呼ばれるクロロフィルを結合している。その機能は光化学系の吸光断面積を拡げるとも、過剰光エネルギー消去のための緩衝帯とも言われるが、PSIとPSIIを簡単に見分けるマーカーとしても有用である。
掲載誌情報
雑誌名: eLife(2023年4月12日付でオンライン出版)
論文タイトル: The photosystem I supercomplex from a primordial green alga Ostreococcus tauri harbors three light-harvesting complex trimers
(原始緑藻Ostreococcus tauriの光化学系I超複合体は3つの三量体集光アンテナ複合体を持つ)
著者:Asako Ishii, Jianyu Shan, Xin Sheng, Eunchul Kim, Akimasa Watanabe, Makio Yokono, Chiyo Noda, Chihong Song, Kazuyoshi Murata, Zhenfeng Liu, Jun Minagawa
DOI:10.7554/eLife.84488 https://elifesciences.org/articles/84488
研究グループ
自然科学研究機構 基礎生物学研究所/生理学研究所/生命創成探究センター/中国科学院生物物理学研究所による国際共同研究の成果です。
研究サポート
本研究は科学研究費補助金「基盤S(課題番号:JP21H05040、研究代表者:皆川純)」、基礎生物学研究所超階層生物学センター・トランスオミクス解析室、モデル植物研究支援施設、生理学研究所共同研究、他のサポートを受けて行われました。
問い合わせ先
研究に関すること
基礎生物学研究所 環境光生物学研究部門
教授 皆川 純(ミナガワ ジュン)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
TEL: 0564-55-7515
E-mail: minagawa_at_nibb.ac.jp
ホームページ: https://www.nibb.ac.jp/photo/
※_at_は@にご変更ください。
報道に関すること
基礎生物学研究所 広報室
TEL: 0564-55-7628 FAX: 0564-55-7597
E-mail: press_at_nibb.ac.jp
生理学研究所 研究力強化戦略室
TEL: 0564-55-7722 FAX: 0564-55-7721
E-mail: pub-adm_at_nips.ac.jp
生命創成探究センター 研究戦略室
TEL: 0564-59-5203 FAX: 0564-59-5202
E-mail: press_at_excells.orion.ac.jp
※_at_は@にご変更ください。