
甲南大学大学院自然科学研究科の久原篤 教授、大西康平 博士研究員(現 広島大学大学院医系科学研究科 博士研究員)、太田茜 特任研究講師、大学共同利用機関法人自然科学研究機構 生命創成探究センター/生理学研究所の曽我部隆彰 准教授、富永真琴 教授らの研究チームは、「ヒトの目で働く光受容体(ロドプシン)などで知られるGタンパク質共役受容体GPCR注1)が神経細胞で温度感知に関わり、体の温度への慣れに関わる」ことを線虫の解析から明らかにしました。
研究チームはシンプルな実験動物である線虫C. エレガンス注2)の温度順化を解析してきました。今回、線虫の温度順化において、温度センサーとして機能するGタンパク質共役受容体GPCRの探索を行いました。遺伝子を抑制する技術であるRNA干渉法注3)を用いて、約1000個のGPCRの中から温度順化に関わるGPCRを探索しました。そして、その中から、温度センサー候補のGPCRとして「SRH-40」を見つけました。
今回見つかったGPCR「SRH-40」を欠損した変異体線虫において、温度への慣れ(温度順化)に異常が見られ、頭部のADL温度受容ニューロン注4)の温度への反応が低下していました。「SRH-40」を味覚ニューロンASE注5)に導入したところ、味覚ニューロンASEが温度に反応するようになりました。同様に、「SRH-40」をショウジョウバエの培養細胞(S2R+細胞注6))に導入したところ、温度への反応が見られました。
以上の結果から、GPCR「SRH-40」が温度を感じる神経細胞で、温度受容に関わり、線虫個体の温度への慣れを調節することが分かりました。これは動物の温度受容神経細胞において、温度受容に関わるGPCRが、体の温度適応を調節することを示す初めてのケースです。感覚の分子機構はヒトから線虫まで保存されていることから、ヒトを含む動物の温度受容機構の理解や、ヒトの温度関連の疾患の原因解明に繋がるのではないかと期待されます。本研究の成果は、2024年2月23日に、英国科学雑誌「Nature communications」 のオンライン速報版で公開されました。
研究の背景と経緯
温度情報は生物が生存していく中で重要な環境因子です。例えば、人間では体温がわずか3℃低下し33℃になると仮死状態になることや、病気の発生率や死亡率は気温変化が大きい時期に増加します。温度が関わる病気は多数あり、急激な温度変化によるヒートショック死や低体温症が社会問題となっており、これらにより亡くなる方の数は交通事故による死亡数よりも多いという報告があります。地球規模でも、温暖化や局所的な寒冷化により生物種の3割が絶滅の危機に瀕すると報告されており、生物の温度適応は世界的な課題となっています。
生物は生育環境から絶え間なく温度情報を受け取り、それに適応することで生存してきました。そのため、生物は温度変化を感知して、それに対応する生体メカニズムを持っています。例えば人間では自律的な体温調節機構が存在し、深部体温と環境温度の変化をそれぞれ感知して情報を統合し、適切な体温調節を行います。線虫からヒトまで多くの動物で進化的に保存された温度センサー分子として、TRP(Transient receptor potential)イオンチャネルが知られており、様々な細胞や組織で発現し、細胞内カルシウム濃度を増加させることにより多くの熱感覚応答に関与しています。
生物の温度応答メカニズムの解明は多方面から望まれている一方で、ヒトの脳は約1000億個の神経細胞で構成されているため、その神経回路は非常に複雑です。そこで本研究チームは、動物の温度応答メカニズムの解明に向けて、ヒトと相同な遺伝子を多く持つシンプルなモデル実験動物である線虫「C.エレガンス」を用いました。C.エレガンスは959個の細胞を持ち、そのうち302個が神経細胞です。
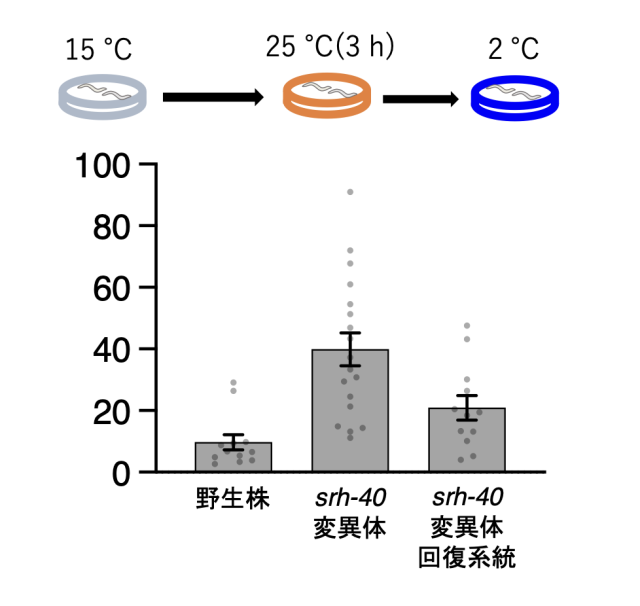
C.エレガンスの温度への適応機構として、これまでに温度順化現象が確認されています。温度順化現象とは、飼育環境の変化に合わせて低温(2℃)への耐性が変化する現象を指します(図1)。具体的には、15℃で飼育した線虫は低温耐性を持ち、2℃に移されても生存可能ですが、25℃で飼育した線虫は低温耐性を持たず、2℃では死滅します(図1)。15℃で飼育した線虫を25℃で3時間以上飼育すると低温耐性を失い、2℃に移されると死滅するようになります。
この低温耐性や温度順化現象を指標に、温度応答に関わる分子や神経細胞の解析が進められてきました。これまでに、低温耐性や温度順化に関わる温度受容ニューロンとして、頭部に位置するASJニューロン注7)とADLニューロンが見つかっています(図2)。これらの温度受容ニューロンで機能する分子はそれぞれ異なります。ASJ温度受容ニューロンでは、環状GMP依存性チャネルTAX-4に依存して温度受容ニューロンの活性化を起こし、シナプスからインスリンを分泌して低温耐性を負に制御します。ADL温度受容ニューロンでは、温度感受性TRPチャネルであるOSM-9とOCR-2が温度受容ニューロンを活性化させます(図2)。
しかし、ASJ温度受容ニューロンでは温度センサー分子が見つかっておらず、ADL温度受容ニューロンでは温度感受性TRPチャネルが発現していますが、その活性はとても弱いものであったため、それ以外の温度センサー分子が存在すると考えられました。
(「研究の背景と経緯」の一部は、過去のプレスリリース「温度への慣れに関わる脳・腸連関 令和4年7月27日 甲南大学」を引用)
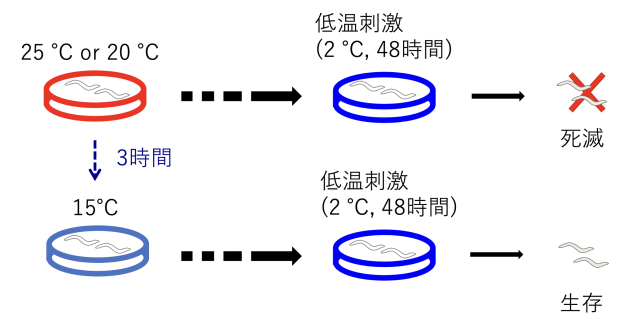
15℃で飼育された線虫は、2℃に置かれても生存できるが(低温耐性)、25℃で飼育された線虫は2℃に置かれると死滅してしまう。15℃で飼育された線虫を25℃で3時間飼育すると低温耐性を失い、死滅する(温度順化)。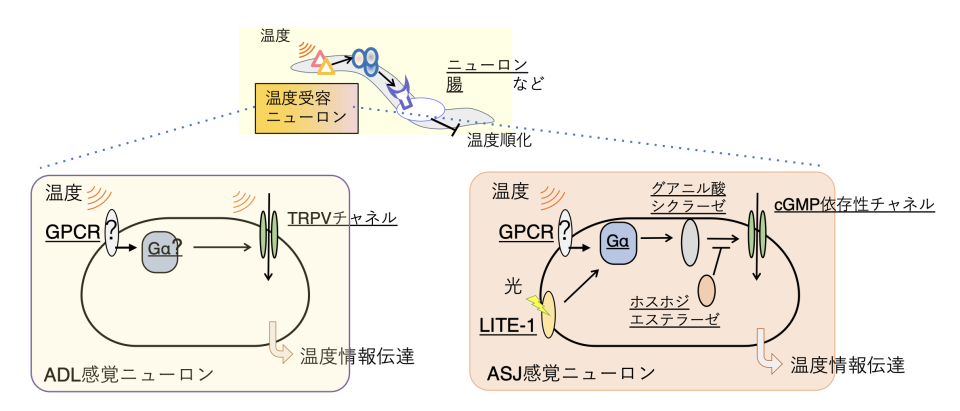
ADL温度受容ニューロンでは、温度感受性TRPVチャネルが温度受容ニューロンの活性化に関わり、ASJ温度受容ニューロンはcGMP依存性チャネルを介して低温耐性を負に制御する。
研究の内容
低温耐性や温度順化に関わるGPCR型温度センサー分子のRNAiスクリーニング
C.エレガンスのASJもしくはADL温度受容ニューロンで機能するGPCR型温度センサー分子の単離するために、RNAiスクリーニングを行いました。C.エレガンスに存在する約1000個のGPCR遺伝子それぞれについてRNA干渉法を用いて遺伝子の機能を抑制して、それらの系統について低温耐性の異常を調べました。この中から、特に強い低温耐性の異常を示したGPCR遺伝子について、更なる解析を行いました(図3)。緑色蛍光タンパク質GFPを用いて約50個のGPCRに関して発現細胞を同定した結果、16個のGPCRがADLもしくはASJ温度受容ニューロンに存在していることを見つけました。次に、これらの中のいくつかのGPCRについて、CRISPR/Cas9法注8)を用いて遺伝子を欠損させた(ノックアウト)個体を作製しました。これは、RNAiにより遺伝子をノックダウンした個体と遺伝子をノックアウトした個体の表現型が異なる場合があるためです。結果として、SRH-40 が欠損した変異体線虫(srh-40変異体)は、15℃から25℃への慣れ(順化)に異常がありました。具体的には、15℃飼育後に25℃で3時間静置したのちに2℃の低温刺激を与えた時に、生存率が上昇する異常が見られました (図4)。
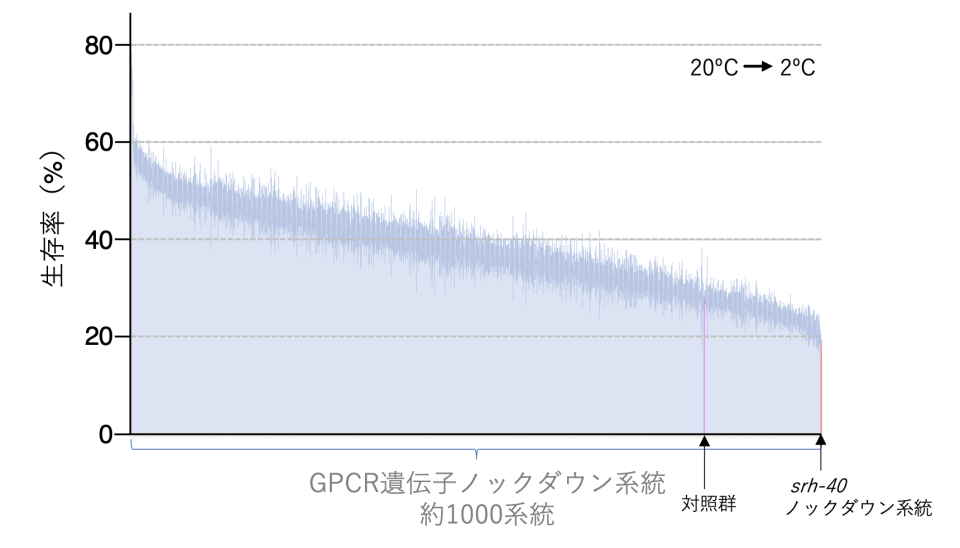
C.エレガンスに存在する約1000個のGPCR遺伝子について1個ずつ遺伝子ノックダウンを行い、低温耐性を調べた。対照系統と比較して低温耐性に異常を示すsrh-40遺伝子などのノックダウン系統を同定した。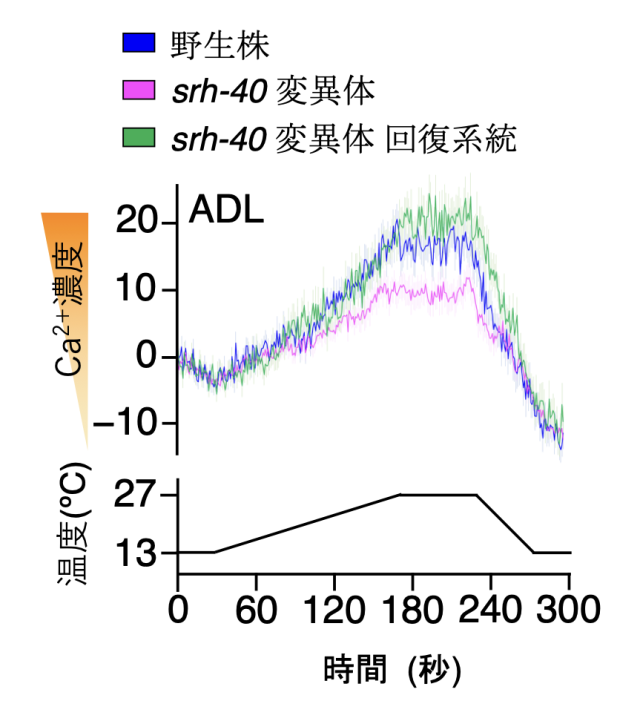
野生株は15℃飼育後に25℃で3時間静置したのちに2℃の低温刺激を与えると死滅するのに対して、srh-40変異体は高い生存率を示しました。また、srh-40変異体の異常は、正常な srh-40遺伝子を導入すると回復しました。
GPCR SRH-40はADL温度受容ニューロンで温度情報伝達に関わる
SRH-40の発現解析を行うとADLニューロンで発現していました。そこでSRH-40がADLニューロンにおいて温度順化を制御するかを調べるために、srh-40 変異体のADLニューロンに正常な srh-40遺伝子を導入したところ、srh-40 変異体の温度順化の異常が回復しました(図4)。
ADLニューロンでにおけるSRH-40が低温耐性に必要にあると判明したため、次に、srh-40 変異体の ADLニューロンの温度刺激に対する神経活動の変化を、光技術を用いたカルシウムイメージング注9)によって調べました。その結果、SRH-40を欠損させた変異体ではADLニューロンの神経活動が低下する異常が見られました (図5)。このsrh-40変異体の異常な神経活性は、 ADLニューロンに正常な srh-40遺伝子を導入すると回復しました (図5)。つまり、SRH-40 が ADLの温度情報伝達に関わることが示唆されました。
srh-40変異体のADLは温度に対する反応が低下する異常を示す。このsrh-40変異体の反応低下は、ADLに正常なsrh-40遺伝子を導入することで回復する。
GPCR SRH-40を介した温度応答
SRH-40 が ADL ニューロンの温度応答に関わっていたため、GPCRである SRH-40が温度を受容しているということを予想しました。この可能性を検証するために、温度上昇には反応しない味覚ニューロン(ASE)にSRH-40を強制発現注10)させ、温度に応答するようになるかをカルシウムイメージング法を用いて調べました。その結果、SRH-40を強制発現させたASE味覚ニューロンは温度上昇に対して応答するようになりました(図6)。
さらに培養細胞(S2R+)にSRH-40とcGMP依存性チャネルを強制発現させ、温度に応答するようになるかをカルシウムイメージング法を用いて調べた結果、SRH-40を強制発現させた培養細胞は温度上昇に対して強く応答するようになりました(図7)。
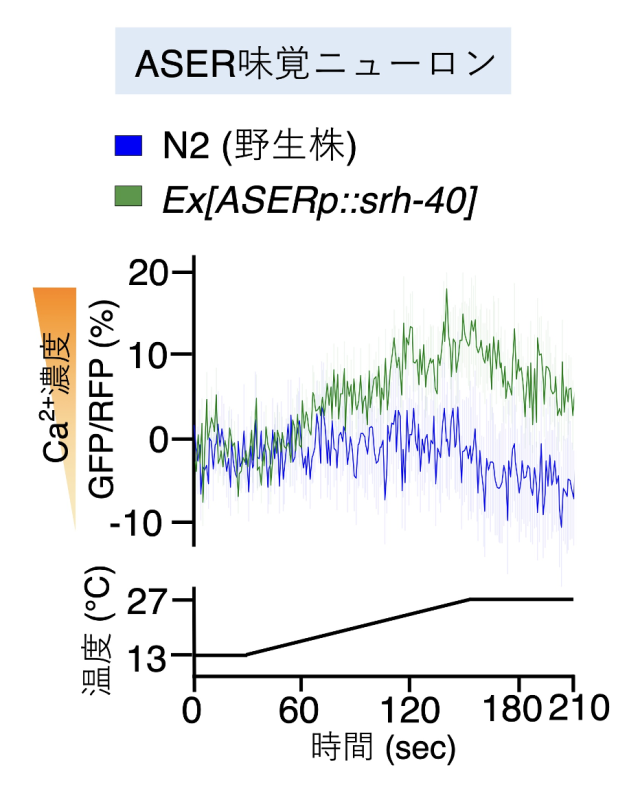
SRH-40を線虫の味覚ニューロンに強制発現させ、温度刺激に対する反応を測定した結果、温度上昇に対してカルシウムイオンの上昇を示した。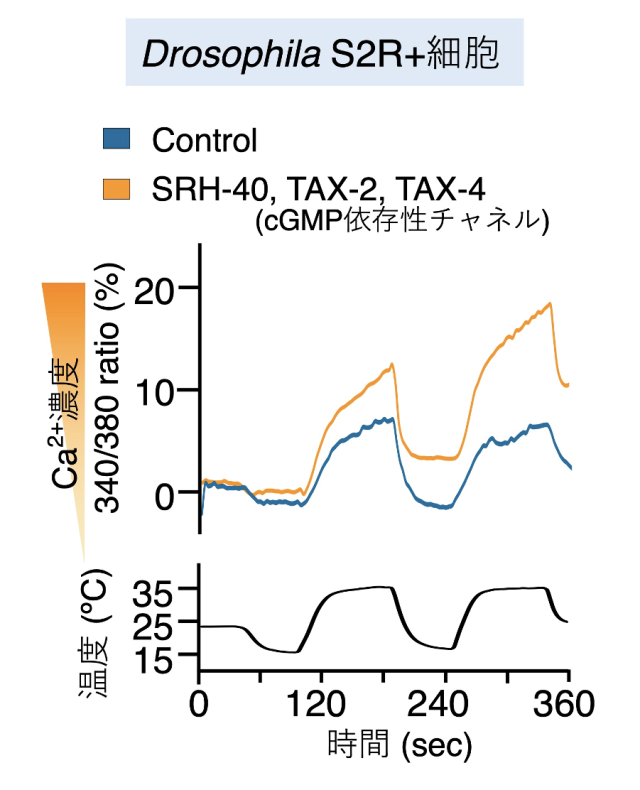
SRH-40を昆虫細胞に強制発現させ、温度に対する反応を測定した結果、SRH-40を発現させた昆虫細胞は温度上昇に対してカルシウムイオンの上昇を示した。
今後の展開
本研究から、線虫からヒトまで保存されている「Gタンパク質共役受容体GPCR」が細胞の温度応答に関わり、線虫では体の温度順化に関わることが見つかりました。この結果は、動物においてGタンパク質共役受容体GPCRが温度受容に関わり個体の温度適応に制御することを示す初めてのケースです。 線虫では、GPCR「SRH-40」を介してADL感覚ニューロンが受け取った温度情報が体の温度順化を調整していると考えられます(図8)。
ヒトにおいて、GPCRであるオプシンが精子の温度への走性に関わることが報告されています。今回見つかったSRH-40の明確なヒトホモログは見つかっていませんが、線虫のGPCR遺伝子にはヒトにも類似遺伝子が存在するものがあるため、今後、ヒトにおける新規の温度受容体GPCRの発見や、温度が関わる未解明の疾患の原因解明に繋がると期待されます。
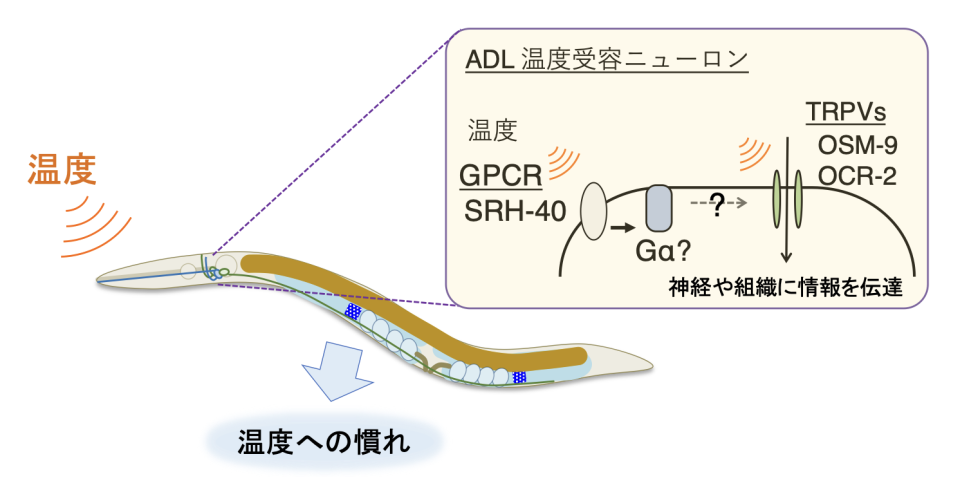
ADLではGPCR SRH-40と温度感受性TRPV OSM-9 OCR-2が温度を受容し、下流で機能する神経や組織などに情報伝達することで個体の温度順化を制御している可能性が考えられる。
研究支援
本研究は、日本医療研究開発機構(AMED)革新的先端研究開発支援事業(PRIME)(23gm6510004h0003、23gm6510014h0002)、日本学術振興会科学研究補助金(23K14235、21H02534、21K19279、22H05512、15H05928、21K06275、21H02531)、甲南学園平生太 郎基金科学研究奨励助成、自然科学研究機構生理学研究所共同利用研究、自然科学研究機構生命創成探究センター共同利用研究、内藤記念科学振興財団、武田科学振興財団、旭硝子財団、光科学技術研究振興財団、島津科学技術振興財団、ブレインサイエンス振興財団、東レ科学振興会の研究費によって行われました。
用語解説
注1)G protein-coupled receptor (GPCR):Gタンパク質共役受容体
生体内に存在する受容体の一つ。7回膜貫通型受容体とも呼ばれ、細胞外の神経伝達物質やホルモンを受けとって、Gタンパク質と呼ばれる三量体タンパクを介して細胞内に情報を伝える。GPCRは線虫からヒトまで保存されており、例えば、目で働く光受容体ロドプシンなどが知られる。
注2)C.エレガンス
正式名称はカエノラブディティス・エレガンス。非寄生性の線虫で、土壌中に生息している。古くから分子遺伝学的解析に使用されており、1998年には多細胞生物で初めて全ゲノムDNAの解読が終了した。ヒトの遺伝子数と同等に約2万個の遺伝子を持ち、類似の遺伝子も多い。
注3)RNA干渉法(RNAi)
遺伝子発現調節技術の一つで、目的遺伝子を標的とした短いRNA配列を発現させてその発現量を低下させることができる。全身あるいは特定の細胞のみで作用させることができる。
注4)ADL(温度受容ニューロン)
エレガンスの頭部に左右一対だけ存在し、環境の感覚情報を受け取る感覚ニューロンで ある。線虫にとって嫌な匂いやフェロモンを感じることができる。最近の研究で、温度も感じることが分かった。
注5)ASE(味覚受容ニューロン)
エレガンスの頭部に左右一対だけ存在し、環境の感覚情報を受け取る感覚ニューロンである。 主に塩味 (NaCI) を受容し、 C. エレガンスが適度な濃度の NaCl に集まる行動に関わっている。
注6)S2R+細胞(キイロショウジョウバエの培養細胞)
S2R+(Schneider 2 Receptor +)細胞は、キイロショウジョウバエDrosophila melanogaster由来の細胞株の1つ。
注7) ASJ(温度受容ニューロン)
エレガンスの頭部に左右一対だけ存在し、環境の感覚情報を受け取る感覚ニューロンで ある。光を感じることができるほか、当研究室の研究から、温度を受容し低温耐性に関わることが分かっている。
注8)CRISPR/Cas9
ゲノム編集技術の一つで、ゲノム配列の任意の部位を自由に編集することができる。二本鎖DNAを切断する酵素Cas9タンパク質と、Cas9が結合する任意のゲノム上のPAM配列(NGG)の上流20 bpの配列を持つガイドRNA(gRNA)を細胞内に導入する。
注9)カルシウムイメージング法
ニューロンの働きに重要である細胞内のカルシウム濃度の変動を色の変化として見る方法 のことを指す。本研究では、カルシウムイオンと結合し、カルシウムの濃度により色が変わる蛍光タンパク質を細胞に導入した。
注10)強制発現
特定の遺伝子を、普段は発現していない細胞に人工的に発現させること。今回、ASER神経細胞において本来は存在しないSRH-40タンパク質を、遺伝子導入により強制的に作らせた。
掲載論文名
“G protein-coupled receptor-based thermosensation determines temperature acclimatization of Caenorhabditis elegans”
(Gタンパク質共役型受容体に基づいた温度受容が線虫の温度順化を決定する)
英国科学雑誌「Nature communications」
DOI: https://doi.org/10.1038/s41467-024-46042-z
発表者
大西 康平(オオニシ コウヘイ)(甲南大学大学院自然科学研究科)
*曽我部 隆彰 (ソカベ タカアキ) (大学共同利用機関法人自然科学研究機構 生命創成探究センター/生理学研究所、准教授)
三浦徹 (ミウラ トオル) (甲南大学理工学部)
富永真琴 (トミナガ マコト) (大学共同利用機関法人自然科学研究機構 生命創成探究センター/生理学研究所、教授)
*太田 茜(オオタ アカネ)(甲南大学理工学部 特任研究講師)
*久原 篤(クハラ アツシ)(甲南大学理工学部/大学院自然科学研究科/統合ニューロバイオロジー研究所、教授)
*責任著者
お問い合わせ先
本研究内容に関すること
 久原 篤(クハラ アツシ)(本研究論文の責任著者)
久原 篤(クハラ アツシ)(本研究論文の責任著者)
甲南大学 理工学部 教授
〒658-8501 兵庫県神戸市東灘区岡本8-9-1 甲南大学理工学部(14号館3F 久原研)
Tel:078-435-2512 Fax:078-435-2539
E-mail: kuhara@konan-u.ac.jp 研究室ホームページ:http://kuharan.com
 大西 康平(オオニシ コウヘイ)(本研究論文の筆頭著者)
大西 康平(オオニシ コウヘイ)(本研究論文の筆頭著者)
広島大学大学院医系科学研究科 生理学及び生物物理学 博士研究員
(研究当時 甲南大学大学院自然科学研究科 博士研究員)
〒734-8553 広島県広島市南区霞一丁目2番3号
Tel:082-257-5122
E-mail: onishi.6.kohei@gmail.com
 曽我部 隆彰(ソカベ タカアキ)(本研究論文の責任著者)
曽我部 隆彰(ソカベ タカアキ)(本研究論文の責任著者)
自然科学研究機構 生命創成探究センター/生理学研究所 准教授
〒444-8787 愛知県岡崎市明大寺町字東山5-1
Tel:0564-59-5287 Fax:0564-59-5285
E-mail: sokabe@nips.ac.jp
 太田 茜(オオタ アカネ)(本研究論文の責任著者)
太田 茜(オオタ アカネ)(本研究論文の責任著者)
甲南大学理工学部 特任研究講師
〒658-8501 兵庫県神戸市東灘区岡本8-9-1 甲南大学理工学部(14号館3F 久原研)
Tel:078-435-2512 Fax:078-435-2539
E-mail: aohta@konan-u.ac.jp
広報に関すること
甲南学園 広報部
〒658-8501 兵庫県神戸市東灘区岡本8-9-1
Tel:078-435-2314 Fax:078-435-2546
E-mail: kouhou@adm.konan-u.ac.jp
自然科学研究機構 生命創成探究センター 研究戦略室
TEL: 0564-59-5203 FAX: 0564-59-5202
E-mail: excells-public@orion.ac.jp
自然科学研究機構 生理学研究所 研究力強化戦略室
TEL: 0564-55-7722 FAX: 0564-55-7721
E-mail: pub-adm@nips.ac.jp