
動物のからだは数多くの細胞から成り立っています。受精卵から秩序正しく動物のからだが形成される上では、個々の細胞がバラバラに振る舞うのではなく、細胞同士が情報をやり取りし合うことが重要です。動物の発生過程では、細胞が分泌する情報分子(分泌性シグナルタンパク質)が、周囲の細胞に受容されることで、細胞の増殖や分化がコントロールされ、組織や器官が秩序正しく形成されていくことが知られています。一方、実際の発生過程においては、一部の細胞の形態が大きく変化することが知られています。このような細胞の形態の変化は細胞相互の位置関係を大きく変える要因になり得ると考えられますが、そのような変化が分泌性シグナルタンパク質を介した細胞間の情報のやり取りにどのような影響を与えるのかということは重要な問題として残されています。特に、分泌性シグナルタンパク質を産生する細胞自身が形態変化を起こした時に、周りの細胞でのシグナルの受容にどのような影響を与えるかということに関してはこれまで明らかになっていませんでした。
自然科学研究機構 生命創成探究センターおよび基礎生物学研究所の篠塚 琢磨 研究員、高田 慎治 教授らの研究グループは、動物の発生過程で重要な分泌性シグナルタンパク質であるWntを産生する細胞に着目し、その形態変化を観察しました。マウスの脊髄神経管の発生過程では、最も背側に位置するroof plate細胞においてWntが産生されます。今回、このroof plate細胞が発生の進行とともに形態変化を起こして長く伸長し、最終的にはその先端が神経前駆細胞に接することを見出しました。さらに、この形態変化にともなってWntを受容する細胞が交替し、伸長したroof plate細胞から分泌されたWntタンパク質が神経前駆細胞に受容されるようになり、細胞増殖を促進することが明らかになりました。このことから、Wntの産生細胞自身が形態変化を起こすことで、遠く離れた細胞にまでWntシグナルが伝達されるという、新たな細胞同士の情報伝達の仕組みが発見されました。本研究成果は2019年1月16日付で発生生物学専門誌のDevelopmentに掲載されました。
研究の背景
細胞間の情報伝達の制御機構の解明は、生物の発生現象を理解するうえで重要な問題です。発生過程における細胞間情報伝達の制御機構を理解するためには、細胞の形態変化や移動によって、周囲の細胞との位置関係が変化し、シグナルの産生細胞と受容細胞の相対的位置が変わることを考慮する必要があります。しかし、このような変化が細胞間シグナル伝達にどのような影響を与えるかということについてはあまり議論がなされてきませんでした。
神経発生の初期のマウスの脊髄神経管では最も背側に位置するroof plate細胞においてWnt1とWnt3aが発現しています。これらのWntは背側神経管の神経前駆細胞の増殖と分化を制御することが知られています。神経発生後期になると、脊髄神経管は内腔の退縮をともなって、その形態を大きく変化させていきます。これまで、ゼブラフィッシュを用いた研究から、神経発生後期にroof plate細胞が腹側へと伸長することが示されていました。このことから、神経管発生におけるroof plate細胞の形態変化は、分泌性シグナルタンパク質の産生細胞の形態変化が、細胞間の情報伝達にどのような影響を与えるかという問題を取り組むのに適した系であると考えられます。しかし、発生後期のroof plateにおいて実際にWntが発現しているのか、また、どこでWntが受容されているのかということについてはこれまで明らかになっていませんでした(図1)。
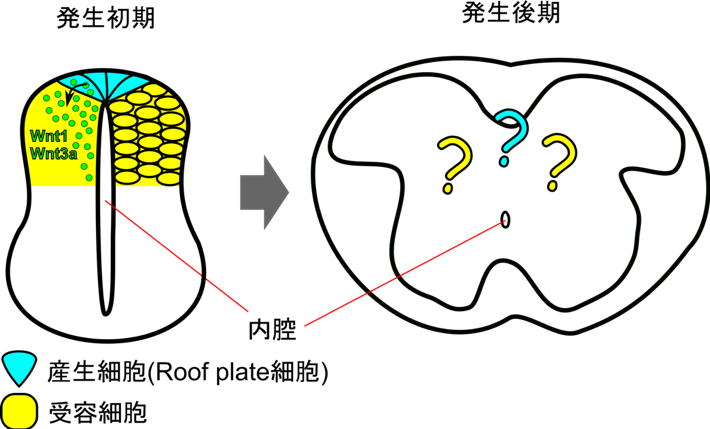
神経発生初期では神経管の最も背側に位置するroof plate細胞(水色)がWnt1 とWnt3aを産生し、その周囲の細胞(黄色)がWntを受容する(左)。神経発生後期になると脊髄神経管は大きく変形し、内腔は退縮する(右)。この変形に伴いroof plate細胞の形はどのように変化するのか、発生後期においてもroof plate細胞はWntを産生し続けるのか、そしてもしそうであるなら、どのような細胞がWntを受容するようになるのかといったことは分かっていなかった。
研究の成果
本研究では、まず、胚発生期のマウスの脊髄神経管を用いて、Wnt産生細胞であるroof plate細胞の形態変化を詳細に調べました。roof plate細胞の一つ一つを観察するために、一部のroof plate細胞のみを標識してみたところ、発生後期になると、それぞれのroof plate細胞は形態変化を起こし、神経管の背側と腹側の両方向に突起を伸長させていることが明らかになりました。さらに、電子顕微鏡による観察から、腹側に伸長した突起の先端は神経管の退縮した内腔の最も背側の部分にまで達していることがわかりました。
そこで、伸長したroof plate細胞においてWntが発現しているのか調べたところ、Wnt1およびWnt3aは確かに発現しており、そのタンパク質は細胞の突起の先端まで運ばれていることがわかりました(図2)。続いて、Wntが受容される細胞を明らかにするため、Wntの情報伝達が起きた細胞を可視化できるマウス(Wntレポーターマウス)を用いて解析を行ったところ、roof plate細胞の突起の先端が接している内腔の周りの細胞がWntを受容していることが明らかになりました。これらの結果から伸長したroof plate細胞で産生されたWntが内腔の周りの細胞で受容され、作用している可能性が考えられました。
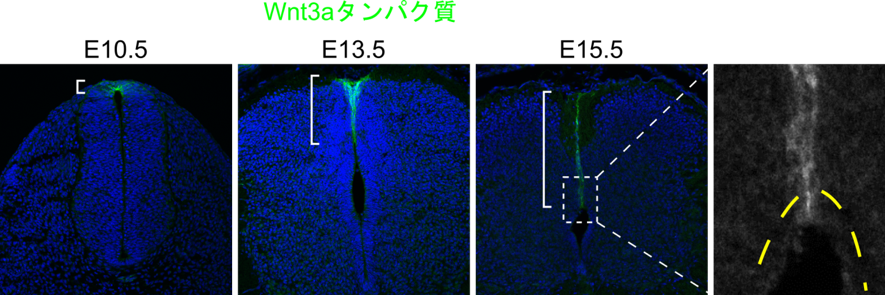
発生にともなってroof plate細胞は腹側へと伸長するが、伸長したroof plate細胞においてもWnt3aタンパク質の発現は観察される。このとき細胞突起の先端までWnt3aタンパク質は輸送されている。
そこで、伸長したroof plate細胞で産生されたWntが内腔の周りの細胞で受容されているかを明らかにするため、伸長したroof plate細胞で特異的にWntタンパク質の分泌を阻害する実験を行いました。roof plate細胞が伸長を開始する受精後13.5日以降にroof plateに特異的にWntタンパク質の分泌に必須の因子であるWlsを欠損させた胚(Wls cKO胚)を解析してみたところ、内腔の周りにおいてのみWntを受容した細胞の数が有意に減少していることがわかりました(図3)。さらに、内腔の周りの細胞は神経前駆細胞を含んでいることが知られているため、これらの細胞の増殖を調べてみたところ、Wls cKO胚では内腔の周りの細胞の細胞増殖も有意に低下していました。これらの結果から、伸長したroof plateから分泌されたWntタンパク質は内腔の周りの細胞に作用し、新たなWntの受容を引き起こすとともに細胞増殖の促進を行っているものと考えられました。
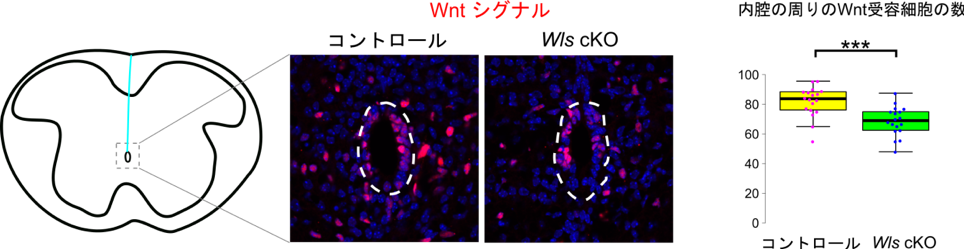
伸長したroof plate細胞からのWntタンパク質の分泌を阻害した胚(Wls cKO胚)では、内腔の周りの細胞で特異的にWntを受容する細胞の数が減少する。
本研究から、発生過程の細胞間情報伝達においては、産生細胞が変形することによって新たな受容細胞との接触が生み出され、遠く離れた細胞にまで情報を伝達することが可能となるという知見が得られました(図4)。
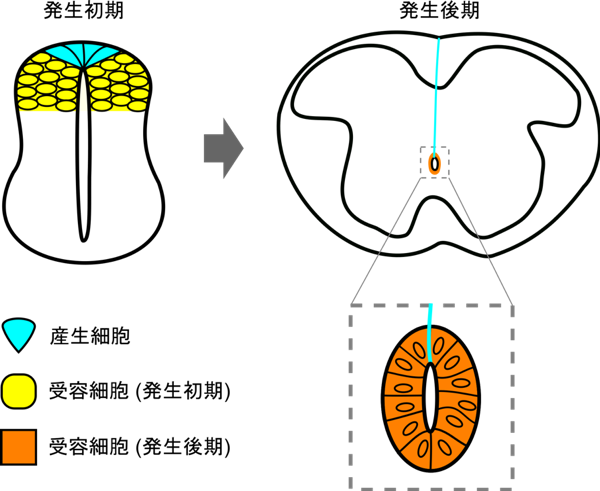
roof plate細胞(水色)は形態変化を起こして背腹軸方向に長く伸長するが、この伸長したroof plate細胞においてもWnt1とWnt3aは引き続き産生される。神経発生初期では背側の神経管の細胞がroof plate細胞が産生するWntを受容していたが(黄色)、roof plate細胞の形態変化にともない、神経発生後期には内腔の周りの細胞がWntを受容するようになる(オレンジ)。
今後の展望
本研究では、脊髄神経管においてWntの産生細胞が変形・伸長して、内腔の周りの神経前駆細胞へとWntの情報伝達を行うことが示されました。Wntの情報は成体の脳においても神経前駆細胞および神経幹細胞を制御することが知られていますが、情報伝達を担うWntの産生細胞は明らかにされていません。脳においても神経前駆細胞および神経幹細胞は、脊髄神経管の内腔から連なる脳室の周りに存在しています。したがって、脳においても脊髄神経管と同様に産生細胞の細胞突起を介した細胞間情報伝達によって神経前駆細胞および神経幹細胞を制御している可能性があり、今後の研究が期待されます。
一方、そもそもroof plate細胞がどのようにして伸長するのか、それがどのように制御されるのかということは神経前駆細胞の増殖制御を考える上でも重要な問題であり、その解明にも興味が持たれます。
掲載誌情報
雑誌名
Development
論文タイトル
Wnt produced by stretched roof-plate cells is required for the promotion of cell proliferation around the central canal of the spinal cord
(伸長した蓋板細胞が産生するWntは脊髄の中心管周囲の細胞増殖の亢進に必要である。)
著者
Takuma Shinozuka, Ritsuko Takada, Shosei Yoshida, Shigenobu Yonemura, and Shinji Takada
DOI
10.1242/dev.159343
研究グループ
本研究は、自然科学研究機構生命創成探究センターおよび基礎生物学研究所(分子発生学研究部門)の篠塚 琢磨 研究員、高田 律子 研究員および高田 慎治 教授らの研究グループを中心として、基礎生物学研究所の吉田 松生 教授(生殖細胞研究部門)、徳島大学大学院医歯薬学研究部および理化学研究所生命機能科学研究センターの米村重信教授からなる共同研究チームにより実施されました。
研究サポート
本研究は、文部科学省科学研究費助成事業および大幸財団の支援を受けて行われました。
発表機関
自然科学研究機構 基礎生物学研究所
自然科学研究機構 生命創成探究センター

お問い合わせ先
自然科学研究機構生命創成探究センター/基礎生物学研究所分子発生学研究部門
教授 高田 慎治(タカダ シンジ)
〒444-8787 愛知県岡崎市明大寺町字東山5-1
TEL: 0564-59-5241
E-mail: stakada_at_nibb.ac.jp ※_at_は@にご変更ください。
ホームページ: http:// www.nibb.ac.jp/cib2/
報道担当
自然科学研究機構 生命創成探究センター 広報担当
TEL: 0564-59-5504
Fax: 0564-59-5226
E-mail: press_at_excells.orion.ac.jp ※_at_は@にご変更ください。
基礎生物学研究所 広報室
TEL: 0564-55-7628
Fax: 0564-55-7597
E-mail: press_at_nibb.ac.jp ※_at_は@にご変更ください。